【論文まとめ】カメの甲羅の多甲(Polyscutellism)|なぜ起きるのか?防げるのか?遺伝するのか?
🐢 カメの甲羅の多甲について|飼育者が最初に知るべき結論
カメの飼育・繁殖を行う中で、
「多甲はなぜ起きるのか?」「遺伝するのか?」「防ぐことはできるのか?」
という疑問は、ほぼ必ず直面するテーマである。
まず、本記事の結論を最初に示す。
多甲は「遺伝」ではなく、「発生初期の環境と発生プログラムが交差して起きる現象」であり、
単一遺伝子で決まる形質ではない。
多甲は、
多遺伝子性素因(polygenic predisposition)と孵卵環境の相互作用によって発生する
多因子性形質(multifactorial trait)である。
🧬 総合概要(結論を先に)
- 多甲は「遺伝病」ではない
- 単一遺伝子によるメンデル遺伝では説明できない
- 発生初期のシグナル異常が直接原因
- 孵卵環境(特に温度・湿度)が最重要因子
- 種親に多甲がいても、環境管理で発生率は低減できる
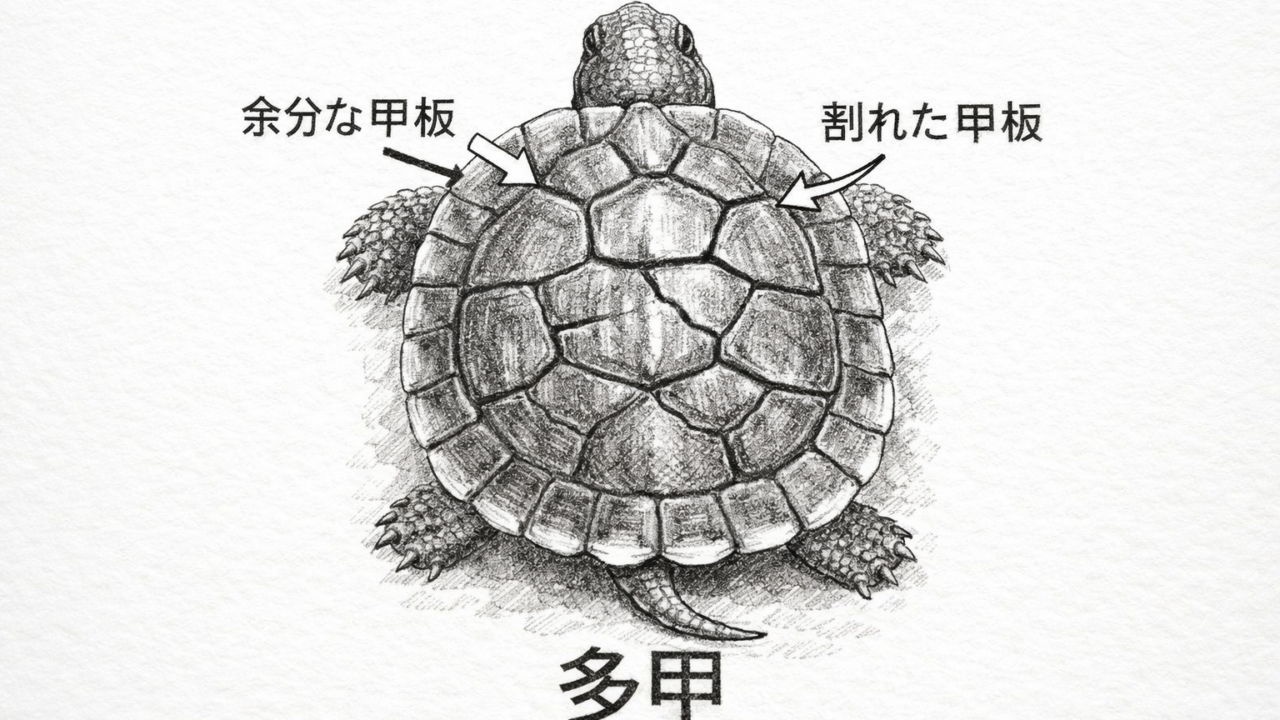
1. 多甲とは何か
多甲(polyscutellism)とは、
カメの甲羅において、本来決まっているスキュート(角質鱗板)の数や配置が乱れ、
通常より多くのスキュートが形成される状態を指す。
これは「スキュート異常(scute anomaly)」の一種であり、
- スキュートの分裂
- 非対称配置
- 融合・欠損
など、さまざまな形で現れる。
形態的には目立つが、
軽度の多甲は直ちに致命的ではないことが多い。
ただし、重度の場合は防御力や成長効率に影響する可能性がある
(McKnight & Ligon, 2014)。
2. 多甲はなぜ起きるのか(発生メカニズム)
2.1 発生初期の分子シグナル異常
カメの甲羅形成は、
胚発生中に出現する カラパシアルリッジ(carapacial ridge) から始まる。
この過程では、
- Wnt
- FGF
- BMP
- Shh(Sonic hedgehog)
といった分子シグナルが、
「どこに、いくつスキュートを作るか」
という空間パターンを精密に制御している
(Moustakas-Verho et al., 2014)。
これらのシグナルは、
濃度・タイミング・相互作用のバランスによって成立しており、
わずかな乱れでもスキュート配置異常が生じる。
最新のトランスクリプトーム解析でも、
Wnt5a と FGF が甲羅形成制御の中核であることが示されている
(Zhang et al., 2021)。
2.2 発生プログラムの確率的ゆらぎ
スキュート配置は、
反応拡散モデル(reaction–diffusion model)で説明できる。
このモデルでは、
- モルフォゲン(化学物質)の濃度勾配
- 微細な確率的揺らぎ
によって、偶然的に異常パターンが生じる。
つまり、多甲は
「発生のミス」ではなく、「発生システムが持つ自然な揺らぎの結果」
としても起こり得る現象である
(Szczygielski et al., 2018)。
3. 多甲は遺伝するのか?
3.1 結論:単純な遺伝形質ではない
多甲は、
- 単一遺伝子
- 優性・劣性
といった メンデル遺伝では説明できない。
研究では、
多遺伝子性素因+環境要因の相互作用として扱われている
(Horváth et al., 2020)。
3.2 遺伝的多様性との関係
遺伝的多様性が低い集団(孤立集団・近交集団)では、
形態異常の頻度が高くなることが報告されている。
ただしこれは、
「多甲が遺伝する」
のではなく、
発生の安定性を支える遺伝的バッファが弱くなる
と解釈される。
4. 環境要因(飼育者が最も介入できる領域)
4.1 孵卵温度・湿度
孵卵中の温度・湿度変動は、
- 胚発生速度
- シグナル伝達タイミング
を乱し、多甲発生率を上昇させる。
特に温度は、
性決定に関与するホルモン経路とも重なっており、
形態形成にも影響を及ぼす
(McKnight & Ligon, 2014)。
4.2 栄養・化学物質
- カルシウム
- ビタミン
- ミネラルバランス
の乱れは、
甲羅の骨化・角質形成に影響する。
また、
環境汚染物質や化学物質も
発生異常の一因となる可能性がある
(Ewert et al., 1984)。
5. 飼育者の実践的結論(重要)
- 多甲の最大要因は「発生初期の環境」
- 産卵後は可能な限り速やかに回収
- 安定した孵卵温度・湿度を維持
- 種親に多甲がいても、孵卵管理次第で発生率は下げられる
つまり、
多甲は「血」ではなく「管理」で左右される
🧩 総合考察
多甲は、
- 発生生物学
- 形態進化
- 環境適応
が交差する、非常に興味深い形質である。
飼育現場においては「欠陥」と見なされがちだが、
学術的には 発生システムの柔軟性を示す現象でもある。
6. 多甲個体は繁殖に使用できるのか
6.1 結論
多甲個体は、一定の条件を満たす場合に限り、繁殖に使用することが可能である。
ただし、その判断は飼育者が発生学的・遺伝学的背景を理解した上で行う必要がある。
多甲は単純な遺伝形質ではなく、
多因子性(発生過程・環境要因・遺伝的素因の相互作用)によって生じる形態異常である。
そのため、
「多甲個体=遺伝的欠陥がある」
「繁殖に使用すべきではない」
といった一律の判断は、
現時点の科学的知見からは支持されていない。
6.2 なぜ一律に「繁殖不可」とは言えないのか
多甲は、以下の理由から
外見的特徴のみで繁殖可否を判断することができない。
① 単一遺伝子形質ではない
多甲はメンデル遺伝形質ではなく、
- 多遺伝子性の遺伝的素因
- 発生初期の環境条件
- 発生プログラムにおける確率的ゆらぎ
が重なって発現する形質である。
そのため、
- 親個体が多甲である
- 子個体も必ず多甲になる
という直接的な因果関係は確認されていない
(Horváth et al., 2020)。
② 環境要因の影響が大きい
同一の親個体を用いた場合であっても、
- 孵卵温度
- 湿度条件
- 温度変動の幅
によって、
多甲の発生頻度は大きく変動することが知られている。
したがって、
多甲の有無そのものよりも、
多甲が生じた要因を説明できるかどうか
が重要となる。
6.3 繁殖に使用すべきでないケース
以下に該当する場合、
多甲個体の繁殖使用は避けるべきである。
重度の多甲や機能障害を伴う個体
- 甲羅の防御機能が著しく低下している
- 成長障害や運動障害が認められる
- 骨格レベルの歪みが疑われる
これらは、単なるスキュート異常を超え、
発生全体に異常が及んでいる可能性を示唆する。
同一系統で多甲が高頻度に継続して発生する場合
- 同一ペア
- 同一孵卵条件
で多甲が連続して発生する場合、
- 遺伝的多様性の低下
- 発生過程の安定性低下
が疑われるため、
繁殖計画や飼育条件の再検討が必要となる。
6.4 条件付きで繁殖使用が可能なケース
以下の条件をすべて満たす場合、
多甲個体を繁殖に使用することは科学的に否定されない。
軽度の多甲で機能障害が認められない個体
- 成長・行動・繁殖に支障がない
- 単一スキュートの分裂など、軽微な形態差に留まる
孵卵環境を安定的かつ高精度に管理できること
- 温度の安定化
- 適切な湿度管理
- 急激な環境変動を回避できる体制
記録と検証を行える体制があること
- 多甲発生率
- 孵化率
- 成長データ
を継続的に記録し、
次世代で評価・検証できる体制が整っていること。
6.5 飼育者に求められる判断基準
多甲個体の繁殖使用において重要なのは、
単なる可否ではなく目的と責任の明確化である。
繁殖が可能であることと、
繁殖を行うべきであることは同義ではない。
- 市場流通を目的とするのか
- 自家検証・記録目的なのか
- 教育・研究目的なのか
目的によって、
許容される判断基準は異なる。
6.6 実践的判断の整理
| 観点 | 繁殖可 | 繁殖不可 |
|---|---|---|
| 多甲の程度 | 軽度・機能正常 | 重度・障害あり |
| 孵卵管理 | 安定・再現性あり | 不安定・属人的 |
| 目的 | 検証・研究 | 無計画な量産 |
| 記録 | 継続的に取得 | 記録なし |
補足まとめ
- 多甲は遺伝形質ではなく、発生過程の結果である
- 繁殖可否は外見ではなく背景要因を基に判断する
- 管理・検証ができない場合は繁殖使用を控えるべきである
- 適切な管理下では、発生理解のための検証対象となり得る
多甲個体の繁殖使用は、
科学的理解と管理責任に基づいて判断されるべき問題である。
✅ まとめ
- 多甲は遺伝病ではない
- 発生初期のシグナル異常が直接原因
- 孵卵環境が最重要因子
- 管理次第で防げる・減らせる
- 多因子性形質として理解すべき
📚 参考文献(APA形式)
- Ewert, M., Firth, S., & Nelson, C. E. (1984). Normal and multiple eggshells in batagurine turtles and their implications for dinosaurs and other reptiles. Canadian Journal of Zoology, 62, 1834–1841.
- Horváth, E., Danko, S., Havas, P., Schindler, M., Šebela, M., Halpern, B., Csibrány, B., Farkas, B., Kaňuch, P., & Uhrin, M. (2020). Variation in shell morphology of the European pond turtle, Emys orbicularis, in fragmented central European populations. Biological Journal of the Linnean Society.
- McKnight, D. T., & Ligon, D. B. (2014). Shell and pattern abnormalities in a population of western chicken turtles. Herpetology.
- Moustakas-Verho, J., et al. (2014). The origin and loss of periodic patterning in the turtle shell. Development, 141, 3033–3039.
- Szczygielski, T., Słowiak, J., & Dróżdż, D. (2018). Shell variability in stem turtles. PeerJ, 6, e6134.
- Zhang, J., et al. (2021). Coordinated WNT and FGF regulate the carapacial ridge development. International Journal of Molecular Sciences, 22(22), 12441.


-320x180.jpeg)
